











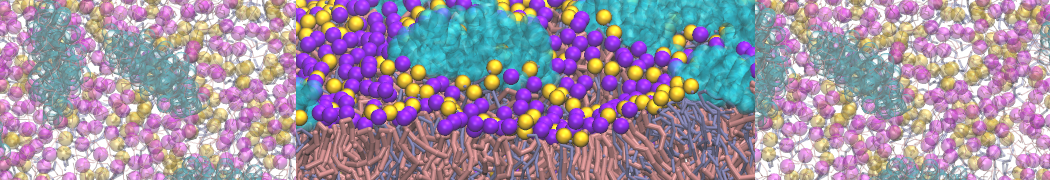
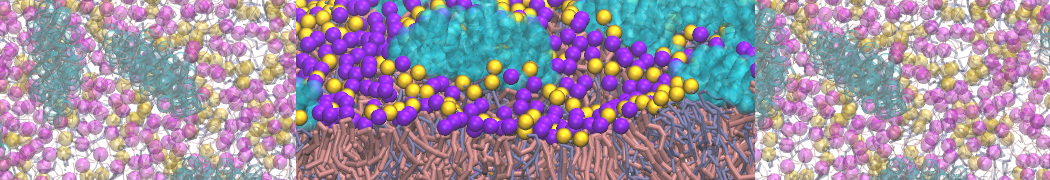
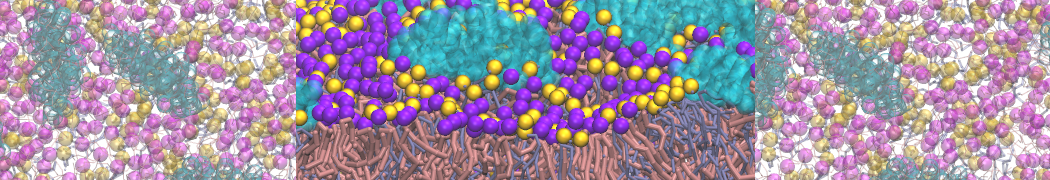
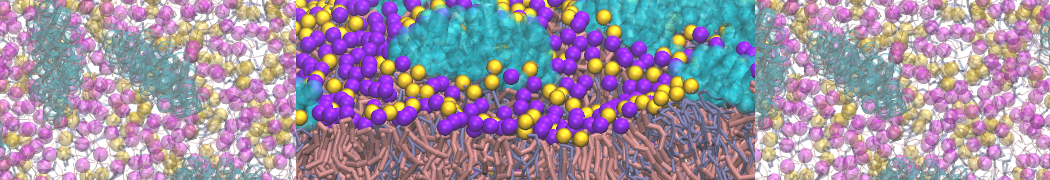






























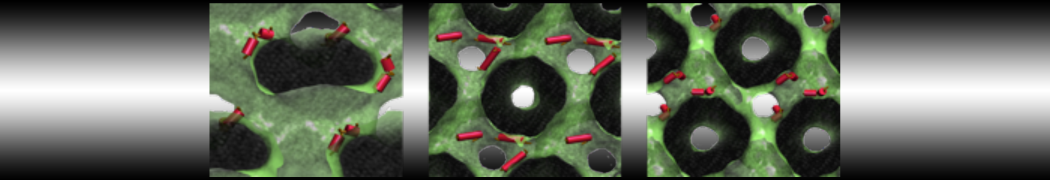
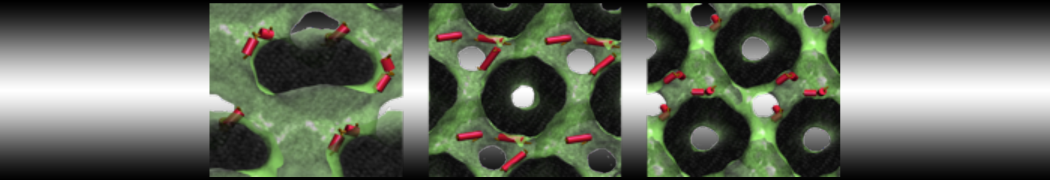
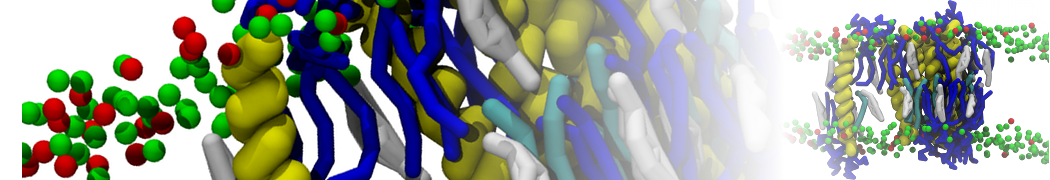
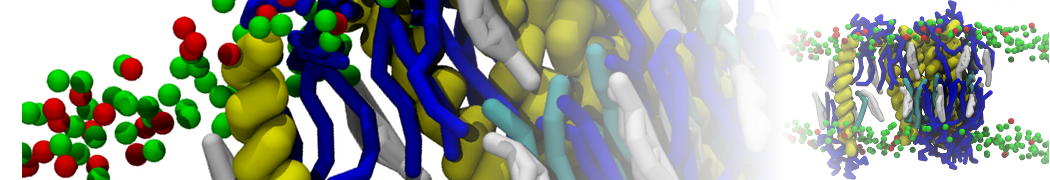
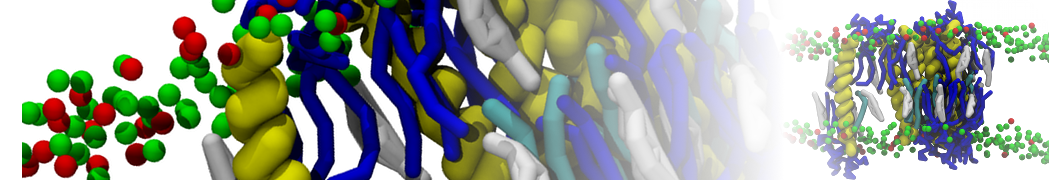
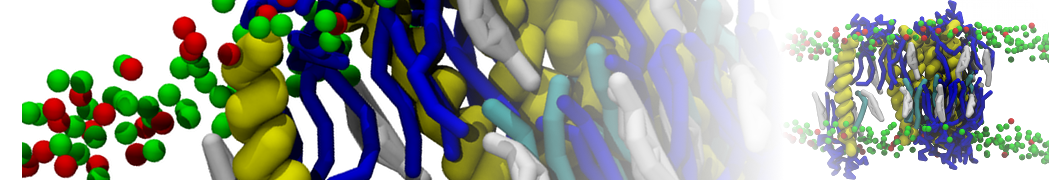
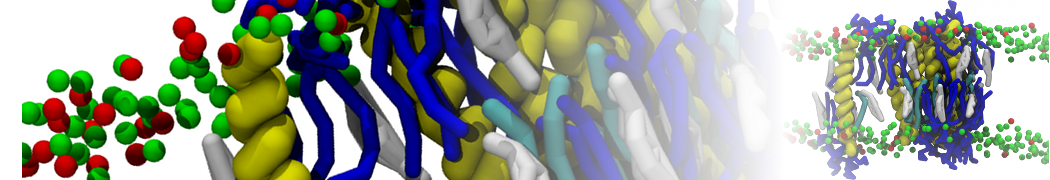
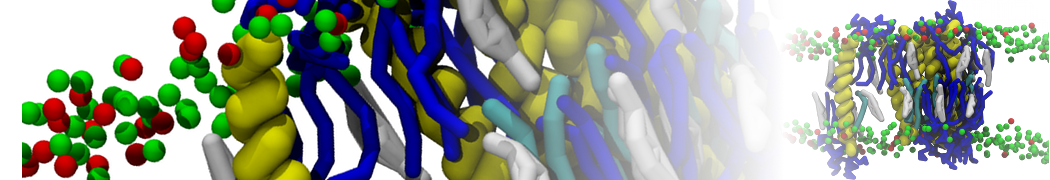
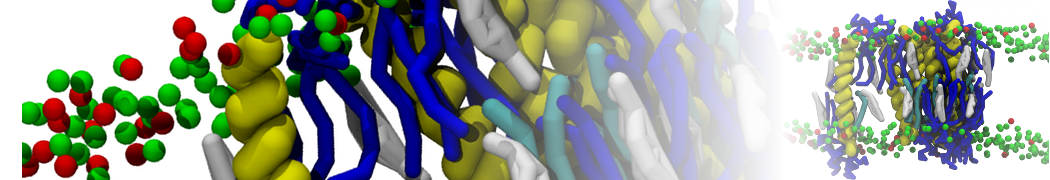
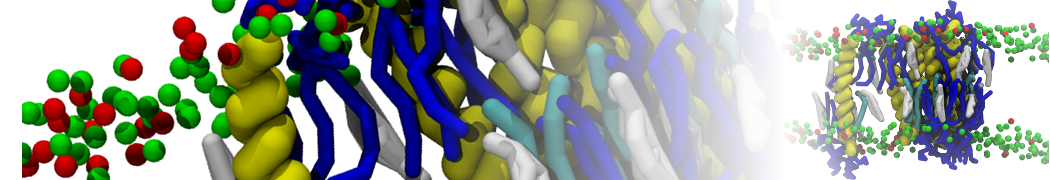
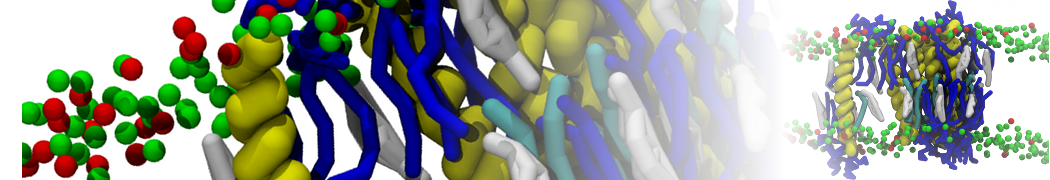
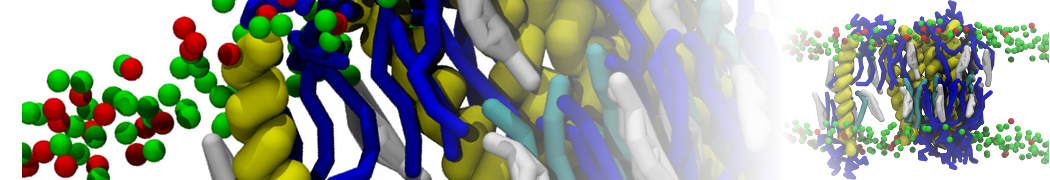











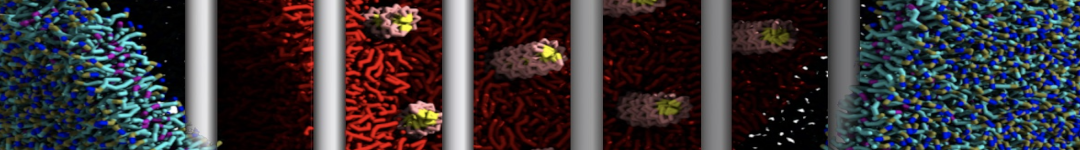
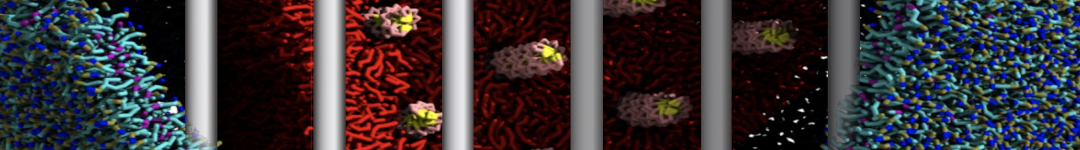
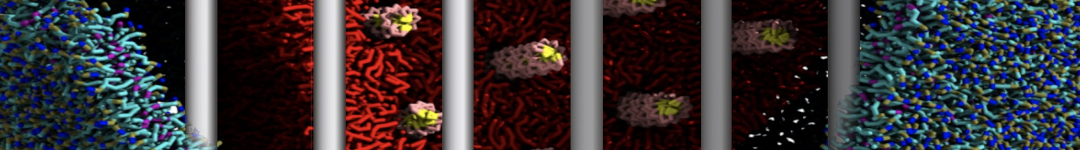
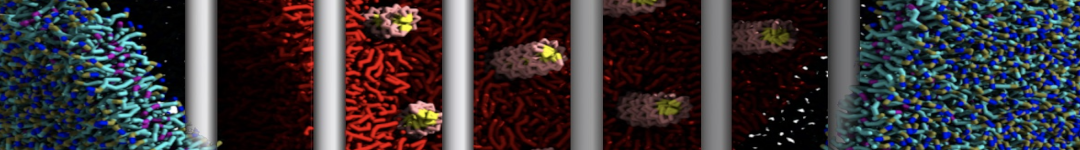






















- Posts: 89
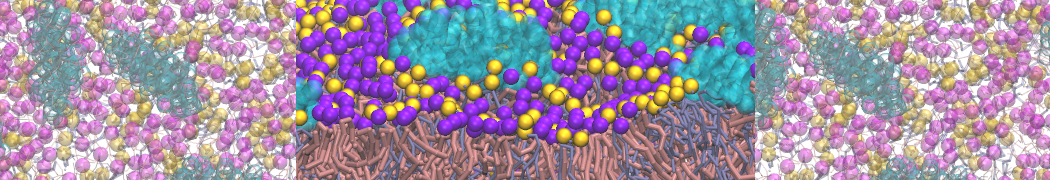
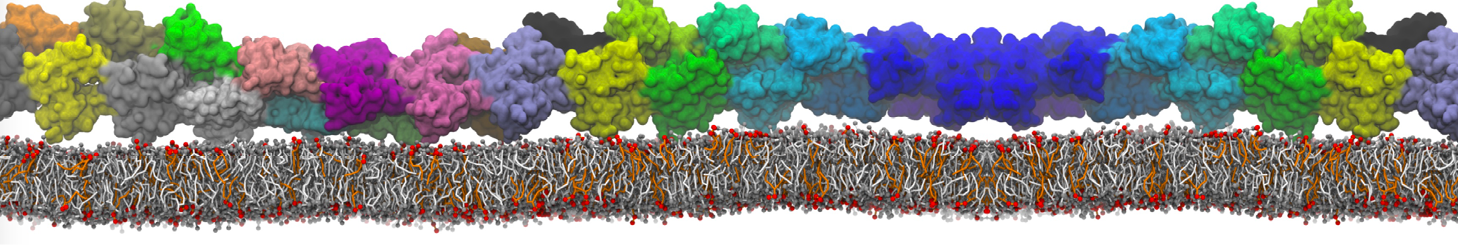
 Pore formation of antimicobial peptide in membrane
Pore formation of antimicobial peptide in membrane
- rongfeng
-
 Topic Author
Topic Author
- Visitor
7 years 9 months ago #5740
by rongfeng
Pore formation of antimicobial peptide in membrane was created by rongfeng
Hi,
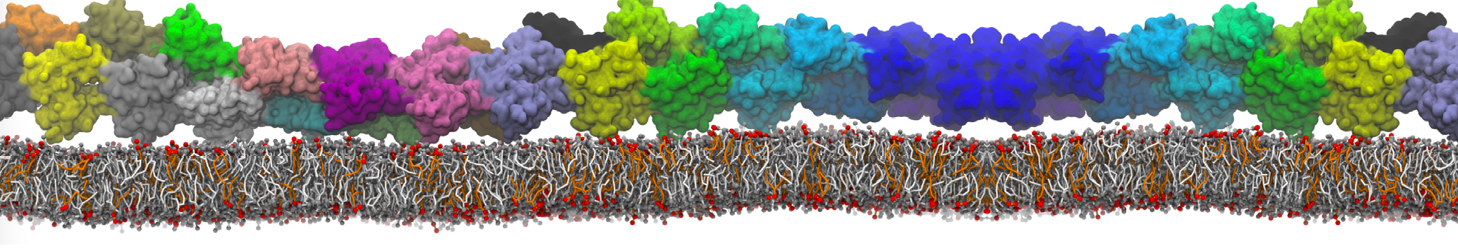
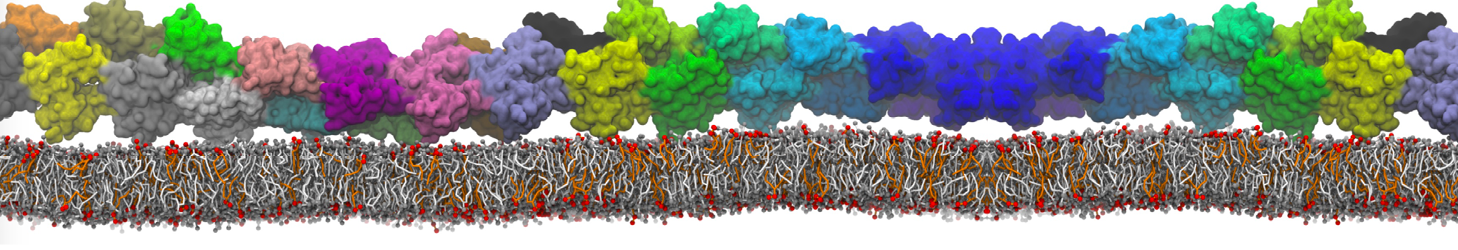
I'm trying to simulate pore formation of Magainin peptide in a POPE/POPG lipid bilayer, but no pore was formed in my simulation. I think maybe there is something wrong with my mdp file.
Here are my mdp files
minim.mdp
integrator = steep
emtol = 1000
emstep = 0.01
nsteps = 50000
nstlist = 1
ns_type = grid
rlist = 1.2
coulombtype = PME
rcoulomb = 1.2
rvdw = 1.2
pbc = xyz
equilibrate.mdp
define = -DPOSRES
integrator = md
tinit = 0.0
dt = 0.020
nsteps = 1000000
nstlog = 10000
nstxtcout = 10000
xtc_precision = 100
nstlist = 10
ns_type = grid
pbc = xyz
rlist = 1.4
cutoff-scheme = group
epsilon_r = 15
coulombtype = Shift
rcoulomb = 1.2
vdw_type = Shift
rvdw_switch = 0.9
rvdw = 1.2
DispCorr = EnerPres
tcoupl = v-rescale
tc-grps = protein membrane solute
tau_t = 1.0 1.0 1.0
ref_t = 310 310 310
; Pressure coupling:
Pcoupl = berendsen
Pcoupltype = semiisotropic
tau_p = 5.0
compressibility = 3e-4 3e-4
ref_p = 1.0 1.0
gen_vel = yes
gen_temp =310
gen_seed = -1
md.mdp
integrator = md
dt = 0.020
nsteps = 15000000000
nstlog = 50000
nstxtcout = 50000
xtc_precision = 100
continuation = yes
nstlist = 20
ns_type = grid
pbc = xyz
rlist = 1.4
cutoff-scheme = group
epsilon_r = 15
coulombtype = Shift
rcoulomb = 1.2
vdw_type = Shift
rvdw_switch = 0.9
rvdw = 1.2
DispCorr = EnerPres
tcoupl = v-rescale
tc-grps = protein membrane solute
tau_t = 1.0 1.0 1.0
ref_t = 310 310 310
Pcoupl = Parrinello-Rahman
Pcoupltype = semiisotropic
tau_p = 12
compressibility = 5e-6 5e-6
ref_p = 1.0 1.0
It will be really helpful if you can give some suggestions.
Thanks!
I'm trying to simulate pore formation of Magainin peptide in a POPE/POPG lipid bilayer, but no pore was formed in my simulation. I think maybe there is something wrong with my mdp file.
Here are my mdp files
minim.mdp
integrator = steep
emtol = 1000
emstep = 0.01
nsteps = 50000
nstlist = 1
ns_type = grid
rlist = 1.2
coulombtype = PME
rcoulomb = 1.2
rvdw = 1.2
pbc = xyz
equilibrate.mdp
define = -DPOSRES
integrator = md
tinit = 0.0
dt = 0.020
nsteps = 1000000
nstlog = 10000
nstxtcout = 10000
xtc_precision = 100
nstlist = 10
ns_type = grid
pbc = xyz
rlist = 1.4
cutoff-scheme = group
epsilon_r = 15
coulombtype = Shift
rcoulomb = 1.2
vdw_type = Shift
rvdw_switch = 0.9
rvdw = 1.2
DispCorr = EnerPres
tcoupl = v-rescale
tc-grps = protein membrane solute
tau_t = 1.0 1.0 1.0
ref_t = 310 310 310
; Pressure coupling:
Pcoupl = berendsen
Pcoupltype = semiisotropic
tau_p = 5.0
compressibility = 3e-4 3e-4
ref_p = 1.0 1.0
gen_vel = yes
gen_temp =310
gen_seed = -1
md.mdp
integrator = md
dt = 0.020
nsteps = 15000000000
nstlog = 50000
nstxtcout = 50000
xtc_precision = 100
continuation = yes
nstlist = 20
ns_type = grid
pbc = xyz
rlist = 1.4
cutoff-scheme = group
epsilon_r = 15
coulombtype = Shift
rcoulomb = 1.2
vdw_type = Shift
rvdw_switch = 0.9
rvdw = 1.2
DispCorr = EnerPres
tcoupl = v-rescale
tc-grps = protein membrane solute
tau_t = 1.0 1.0 1.0
ref_t = 310 310 310
Pcoupl = Parrinello-Rahman
Pcoupltype = semiisotropic
tau_p = 12
compressibility = 5e-6 5e-6
ref_p = 1.0 1.0
It will be really helpful if you can give some suggestions.
Thanks!
Please Log in or Create an account to join the conversation.
- mnmelo
-
- Offline
- Admin
Less
More
7 years 8 months ago #5764
by mnmelo
Replied by mnmelo on topic Pore formation of antimicobial peptide in membrane
Hi Rongfeng,
Pore formation is notoriously tricky with Martini. It appears that the energy of Martini pore-formation is too high , at least compared to atomistic behavior.
In the past pore formation has been observed using polarizable water and PME electrostatics, if the peptides are placed only on one side of the bilayer. However, this seems driven more by the direct and almost unshielded charge interactions with the PG headgroups of the opposing leaflet, and by a tendency of PME to cause such a system to expand in the XY-plane. Also, adding physiological amounts of counterions (~150mM) abolishes this pore formation.
I'm sorry I can't be of more help,
Manel
Pore formation is notoriously tricky with Martini. It appears that the energy of Martini pore-formation is too high , at least compared to atomistic behavior.
In the past pore formation has been observed using polarizable water and PME electrostatics, if the peptides are placed only on one side of the bilayer. However, this seems driven more by the direct and almost unshielded charge interactions with the PG headgroups of the opposing leaflet, and by a tendency of PME to cause such a system to expand in the XY-plane. Also, adding physiological amounts of counterions (~150mM) abolishes this pore formation.
I'm sorry I can't be of more help,
Manel
Please Log in or Create an account to join the conversation.
- roshanak
-
- Offline
- Junior Boarder
Less
More
- Posts: 30
7 years 2 days ago - 7 years 2 days ago #7235
by roshanak
Replied by roshanak on topic Pore formation of antimicobial peptide in membrane
Hi all
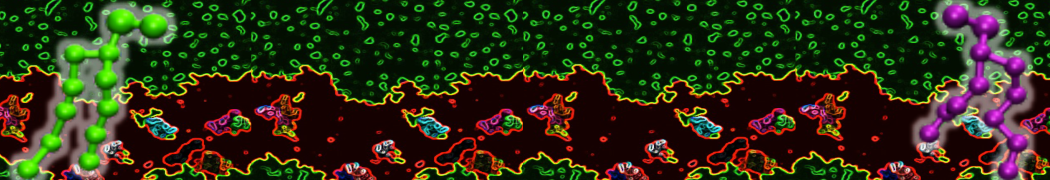
I have similar question. I'm simulating some AMP in center of membrane. I've made a pore via arrangement of peptides. when I ran MD, after a few ns, pore closed.
I've tried martini 2.2p force field with PME and polarizable water. also martini 2.2 and usual water.
I studied some article about membrane poration via antimicrobial peptides. but I didn't find my solution.
would you please get me some hints?
thank you.
I have similar question. I'm simulating some AMP in center of membrane. I've made a pore via arrangement of peptides. when I ran MD, after a few ns, pore closed.
I've tried martini 2.2p force field with PME and polarizable water. also martini 2.2 and usual water.
I studied some article about membrane poration via antimicrobial peptides. but I didn't find my solution.
would you please get me some hints?
thank you.
Last edit: 7 years 2 days ago by roshanak.
Please Log in or Create an account to join the conversation.
- roshanak
-
- Offline
- Junior Boarder
Less
More
- Posts: 30
6 years 11 months ago - 6 years 11 months ago #7263
by roshanak
Replied by roshanak on topic Pore formation of antimicobial peptide in membrane
I need to help ,so much. would you please guide me?
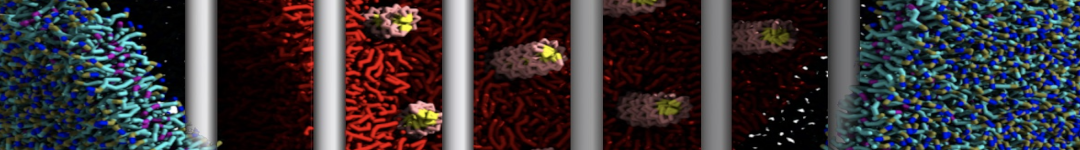
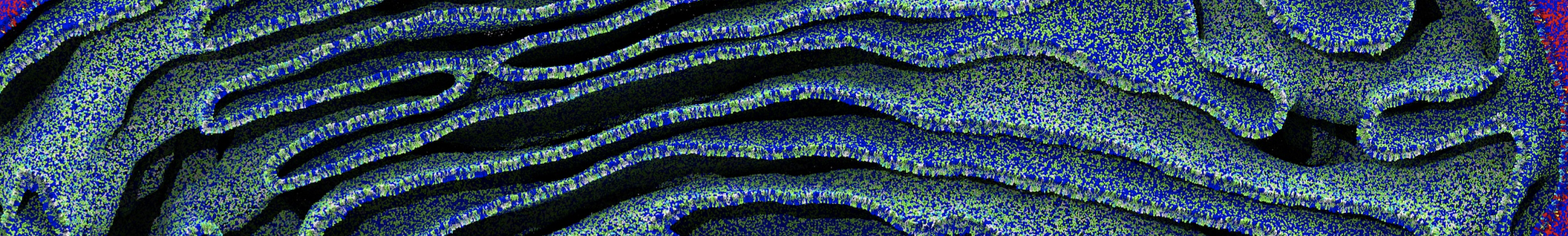
My peptides haven't penetrated and their pore hasn't remain. but with all atom scale, these have happened.
My peptides haven't penetrated and their pore hasn't remain. but with all atom scale, these have happened.
Last edit: 6 years 11 months ago by roshanak.
Please Log in or Create an account to join the conversation.
- Pim
-
- Offline
- Expert Boarder
Less
More
- Posts: 105
6 years 11 months ago #7266
by Pim
Replied by Pim on topic Pore formation of antimicobial peptide in membrane
I'm assuming that in real life this peptide DOES form pores? It might be that your at the limits of what Martini can do for you if pores are collapsing spontaneously. One last trick that might work: one of my colleagues here in Groningen managed to find pore formation for an AMP by two methods:
1) preventing buckling of the membrane by restricting headgroup movement in the out-of-membrane-plane direction, i.e. position restraining the z coordinate of lipid heads
2) because restraints are a strong bias, the same was also achieved using the less biasing flat-bottom potential (requires gromacs 5) on the head-group z coordinate, allowing the lipids to only be inside the initial membrane plane and not above/below.
Good luck,
Pim
1) preventing buckling of the membrane by restricting headgroup movement in the out-of-membrane-plane direction, i.e. position restraining the z coordinate of lipid heads
2) because restraints are a strong bias, the same was also achieved using the less biasing flat-bottom potential (requires gromacs 5) on the head-group z coordinate, allowing the lipids to only be inside the initial membrane plane and not above/below.
Good luck,
Pim
Please Log in or Create an account to join the conversation.
- roshanak
-
- Offline
- Junior Boarder
Less
More
- Posts: 30
6 years 11 months ago #7274
by roshanak
Replied by roshanak on topic Pore formation of antimicobial peptide in membrane
Thanks for reply. I must try it.
would you please recommend some article about similar problem? (I see list of articles in martini site and study some of them and I find some articles with search in google too. but I think that I need more than this.)
would you please recommend some article about similar problem? (I see list of articles in martini site and study some of them and I find some articles with search in google too. but I think that I need more than this.)
Please Log in or Create an account to join the conversation.
- roshanak
-
- Offline
- Junior Boarder
Less
More
- Posts: 30
6 years 11 months ago #7278
by roshanak
Replied by roshanak on topic Pore formation of antimicobial peptide in membrane
Dear Pim, I must add some description, I put my peptides in center of membrane, and I made a pore by arranging peptides. In this time, head group of lipids buckled to pore and pore was open and full of water. after a few ns the peptides aggregated and went up, near surface of membrane, In this time pore got close (disappear) and head groups of lipid came back to membrane plane.
Please Log in or Create an account to join the conversation.
- Pim
-
- Offline
- Expert Boarder
Less
More
- Posts: 105
6 years 11 months ago #7280
by Pim
Replied by Pim on topic Pore formation of antimicobial peptide in membrane
Yeah, I understood that from your previous posts. It's a bit worrying that you don't just not see pore formation, but that the pore collapses and the peptides are even expelled from the membrane if you start from a pore structure. That's the reasoning behind saying that you might be at the limit of martini, somehow the representation of your peptide, pore or membrane is not good enough. The suggestions of using flat-bottom potentials / restraints is therefore also not guaranteed to work. We have not published any papers using them yet as far as I know, the manuscript is still in preparation.
Please Log in or Create an account to join the conversation.
- roshanak
-
- Offline
- Junior Boarder
Less
More
- Posts: 30
6 years 11 months ago - 6 years 11 months ago #7281
by roshanak
Replied by roshanak on topic Pore formation of antimicobial peptide in membrane
ok thanks for reply.
I hope that solution be found :)
Is it possible that I made mistake in some options in mdp files?
Could you please check my files ?
minimization.mdp
;define = -DFLEXIBLE
define = -DPOSRES
integrator = steep algorithm
dt = 0.02
nsteps = 50000
nstcomm = 100
comm-grps =
nstxout = 5000
nstvout = 5000
nstfout = 0
nstlog = 1000
nstenergy = 1000
nstxtcout = 3000
xtc_precision = 100
xtc-grps =
energygrps = System
nstlist = 10
ns_type = grid
pbc = xyz
rlist = 1.4
coulombtype = PME
pme_order = 4 ; cubic interpolation
fourierspacing = 0.16 ; grid spacing for FFT
rcoulomb = 1.4
epsilon_r = 2.5 ; 2.5 (with polarizable water)
vdw_type = Shift
;vdw_modifier =Force-switch
rvdw_switch = 0.9
rvdw = 1.2
ewald-rtol = 1e-5
;cutoff-scheme = group
;coulomb-modifier = Potential-shift-verlet
;epsilon_rf = 0
;verlet-buffer-tolerance = 0.005
constraints = none
constraint_algorithm = Lincs
continuation = no
lincs_order = 8
lincs_warnangle = 30
equilibration.mdp
; STANDARD MD INPUT OPTIONS FOR MARTINI 2.x
; Updated 02 feb 2013 by DdJ
;
; for use with GROMACS 4.5/4.6
;
title = Martini
define = -DPOSRES
;define = -DFLEXIBLE
integrator = md
dt = 0.02
nsteps = 500000
nstcomm = 100
comm-grps = Protein_DPPC PW_ION
nstxout = 0
nstvout = 0
nstfout = 0
nstlog = 1000
nstenergy = 100
nstxtcout = 3000
xtc_precision = 100
xtc-grps =
energygrps = Protein_DPPC PW_ION
nstlist = 10
ns_type = grid
pbc = xyz
rlist = 1.4
coulombtype = PME pairlist) ;PME
pme_order = 4
fourierspacing = 0.16
rcoulomb = 1.4
epsilon_r = 2.5
vdw_type = Shift
rvdw_switch = 0.9
rvdw = 1.2
;vdw_modifier = None
ewald-rtol = 1e-5
;cutoff-scheme = group
;coulomb-modifier = None ;Potential-shift-verlet
;epsilon_rf = 0 ; epsilon_rf = 0 really means epsilon_rf = infinity
;verlet-buffer-tolerance = 0.005
tcoupl = v-rescale
tc-grps = Protein DPPC PW_ION
tau_t = 1.0 1.0 1.0
ref_t = 323 323 323
Pcoupl = berendsen ; ; parrinello-rahman ; parrinello-rahman
Pcoupltype = semiisotropic
tau_p = 12 ; 12.0 12.0 ;parrinello-rahman is more stable with larger tau-p, DdJ, 20130422
compressibility = 3e-4 3e-4 ; 3e-4
ref_p = 1.0 1.0 ; 1.0 1.0
refcoord-scaling =com
gen_vel = no
gen_temp = 320
gen_seed = 473529
constraints = none
constraint_algorithm = Lincs
continuation = no
lincs_order = 8
lincs_warnangle = 30
md.mdp
; STANDARD MD INPUT OPTIONS FOR MARTINI 2.x
; Updated 02 feb 2013 by DdJ
;
; for use with GROMACS 4.5/4.6
title = Martini
integrator = md
dt = 0.02
nsteps = 50000000
nstcomm = 100
comm-grps = Protein_DPPC PW_ION
nstxout = 0
nstvout = 0
nstfout = 0
nstlog = 10000
nstenergy = 100
nstxtcout = 5000
xtc_precision = 100
xtc-grps =
energygrps = Protein DPPC PW_ION
nstlist = 10
ns_type = grid
pbc = xyz
rlist = 1.4
coulombtype = PME
pme_order = 4
fourierspacing = 0.16
rcoulomb = 1.4
epsilon_r = 2.5
vdw_type = Shift
rvdw_switch = 0.9
rvdw = 1.2
;vdw_modifier = None
ewald-rtol = 1e-5
;cutoff-scheme = group
;coulomb-modifier = Potential-shift
;vdw-modifier = Potential-shift
;epsilon_rf = 0 ; epsilon_rf = 0 really means epsilon_rf = infinity
;verlet-buffer-drift = 0.005
tcoupl = Nose-Hoover
tc-grps = Protein DPPC PW_ION
tau_t = 6.0 6.0 6.0
ref_t = 323 323 323
Pcoupl = parrinello-rahman ;berendsen ; parrinello-rahman ; parrinello-rahman
Pcoupltype = semiisotropic ;isotropic
tau_p = 12.0 ; 12.0 12.0 ;parrinello-rahman is more stable with larger tau-p, DdJ, 20130422
compressibility = 3e-4 3e-4 ; 3e-4
ref_p = 1.0 1.0 ; 1.0 1.0
gen_vel = no
gen_temp = 320
gen_seed = 473529
constraints = none
constraint_algorithm = Lincs
continuation = no
lincs_order = 8
lincs_warnangle = 30
I hope that solution be found :)
Is it possible that I made mistake in some options in mdp files?
Could you please check my files ?
minimization.mdp
;define = -DFLEXIBLE
define = -DPOSRES
integrator = steep algorithm
dt = 0.02
nsteps = 50000
nstcomm = 100
comm-grps =
nstxout = 5000
nstvout = 5000
nstfout = 0
nstlog = 1000
nstenergy = 1000
nstxtcout = 3000
xtc_precision = 100
xtc-grps =
energygrps = System
nstlist = 10
ns_type = grid
pbc = xyz
rlist = 1.4
coulombtype = PME
pme_order = 4 ; cubic interpolation
fourierspacing = 0.16 ; grid spacing for FFT
rcoulomb = 1.4
epsilon_r = 2.5 ; 2.5 (with polarizable water)
vdw_type = Shift
;vdw_modifier =Force-switch
rvdw_switch = 0.9
rvdw = 1.2
ewald-rtol = 1e-5
;cutoff-scheme = group
;coulomb-modifier = Potential-shift-verlet
;epsilon_rf = 0
;verlet-buffer-tolerance = 0.005
constraints = none
constraint_algorithm = Lincs
continuation = no
lincs_order = 8
lincs_warnangle = 30
equilibration.mdp
; STANDARD MD INPUT OPTIONS FOR MARTINI 2.x
; Updated 02 feb 2013 by DdJ
;
; for use with GROMACS 4.5/4.6
;
title = Martini
define = -DPOSRES
;define = -DFLEXIBLE
integrator = md
dt = 0.02
nsteps = 500000
nstcomm = 100
comm-grps = Protein_DPPC PW_ION
nstxout = 0
nstvout = 0
nstfout = 0
nstlog = 1000
nstenergy = 100
nstxtcout = 3000
xtc_precision = 100
xtc-grps =
energygrps = Protein_DPPC PW_ION
nstlist = 10
ns_type = grid
pbc = xyz
rlist = 1.4
coulombtype = PME pairlist) ;PME
pme_order = 4
fourierspacing = 0.16
rcoulomb = 1.4
epsilon_r = 2.5
vdw_type = Shift
rvdw_switch = 0.9
rvdw = 1.2
;vdw_modifier = None
ewald-rtol = 1e-5
;cutoff-scheme = group
;coulomb-modifier = None ;Potential-shift-verlet
;epsilon_rf = 0 ; epsilon_rf = 0 really means epsilon_rf = infinity
;verlet-buffer-tolerance = 0.005
tcoupl = v-rescale
tc-grps = Protein DPPC PW_ION
tau_t = 1.0 1.0 1.0
ref_t = 323 323 323
Pcoupl = berendsen ; ; parrinello-rahman ; parrinello-rahman
Pcoupltype = semiisotropic
tau_p = 12 ; 12.0 12.0 ;parrinello-rahman is more stable with larger tau-p, DdJ, 20130422
compressibility = 3e-4 3e-4 ; 3e-4
ref_p = 1.0 1.0 ; 1.0 1.0
refcoord-scaling =com
gen_vel = no
gen_temp = 320
gen_seed = 473529
constraints = none
constraint_algorithm = Lincs
continuation = no
lincs_order = 8
lincs_warnangle = 30
md.mdp
; STANDARD MD INPUT OPTIONS FOR MARTINI 2.x
; Updated 02 feb 2013 by DdJ
;
; for use with GROMACS 4.5/4.6
title = Martini
integrator = md
dt = 0.02
nsteps = 50000000
nstcomm = 100
comm-grps = Protein_DPPC PW_ION
nstxout = 0
nstvout = 0
nstfout = 0
nstlog = 10000
nstenergy = 100
nstxtcout = 5000
xtc_precision = 100
xtc-grps =
energygrps = Protein DPPC PW_ION
nstlist = 10
ns_type = grid
pbc = xyz
rlist = 1.4
coulombtype = PME
pme_order = 4
fourierspacing = 0.16
rcoulomb = 1.4
epsilon_r = 2.5
vdw_type = Shift
rvdw_switch = 0.9
rvdw = 1.2
;vdw_modifier = None
ewald-rtol = 1e-5
;cutoff-scheme = group
;coulomb-modifier = Potential-shift
;vdw-modifier = Potential-shift
;epsilon_rf = 0 ; epsilon_rf = 0 really means epsilon_rf = infinity
;verlet-buffer-drift = 0.005
tcoupl = Nose-Hoover
tc-grps = Protein DPPC PW_ION
tau_t = 6.0 6.0 6.0
ref_t = 323 323 323
Pcoupl = parrinello-rahman ;berendsen ; parrinello-rahman ; parrinello-rahman
Pcoupltype = semiisotropic ;isotropic
tau_p = 12.0 ; 12.0 12.0 ;parrinello-rahman is more stable with larger tau-p, DdJ, 20130422
compressibility = 3e-4 3e-4 ; 3e-4
ref_p = 1.0 1.0 ; 1.0 1.0
gen_vel = no
gen_temp = 320
gen_seed = 473529
constraints = none
constraint_algorithm = Lincs
continuation = no
lincs_order = 8
lincs_warnangle = 30
Last edit: 6 years 11 months ago by roshanak.
Please Log in or Create an account to join the conversation.
Time to create page: 0.103 seconds