




















































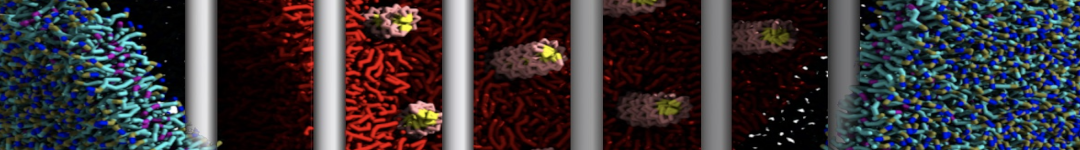






















- Posts: 21
 Martini studies on aggregation of Amino acids in Lipid bilayer
Martini studies on aggregation of Amino acids in Lipid bilayer
- shakira
-
 Topic Author
Topic Author
- Offline
- Junior Boarder
Less
More
3 years 7 months ago #8683
by shakira
Martini studies on aggregation of Amino acids in Lipid bilayer was created by shakira
Hi all
Can anyone guide me to find MARTINI based papers addressing the aggregation behaviour of Aminoacids inside lipid bilayer like simple DPPC. We have seen aminoacids like PHE aggregates inside lipid bilayer from experimental papers. So we need to assess whether Martini FF can be used to address such problems. Are similar works previously done with Martini. I could see papers for peptide aggregation but not simple AA.
Thanks in advance
Can anyone guide me to find MARTINI based papers addressing the aggregation behaviour of Aminoacids inside lipid bilayer like simple DPPC. We have seen aminoacids like PHE aggregates inside lipid bilayer from experimental papers. So we need to assess whether Martini FF can be used to address such problems. Are similar works previously done with Martini. I could see papers for peptide aggregation but not simple AA.
Thanks in advance
Please Log in or Create an account to join the conversation.
- shakira
-
Topic Author
- Offline
- Junior Boarder
Less
More
- Posts: 21
3 years 7 months ago #8690
by shakira
Replied by shakira on topic Martini studies on aggregation of Amino acids in Lipid bilayer
Hi all
Considering many of the limitations of coarse grained Martini water, I guess we may not be able to see the aggregation of hydrophobic amino acids like Phenylalanine (PHE) in bulk water or at the lipid membrane surface. As this paper ( pubs.rsc.org/ko/content/articlehtml/2013/cs/c3cs60093a ) suggests water ordering and strong water interactions are challenging to be reproduced by CG water.
Thus we many not be able to see such aggregated structures for PHE like in all-atom simulations right?
Any guidance on this query will be hugely helpful.
Thanks in advance
Considering many of the limitations of coarse grained Martini water, I guess we may not be able to see the aggregation of hydrophobic amino acids like Phenylalanine (PHE) in bulk water or at the lipid membrane surface. As this paper ( pubs.rsc.org/ko/content/articlehtml/2013/cs/c3cs60093a ) suggests water ordering and strong water interactions are challenging to be reproduced by CG water.
Thus we many not be able to see such aggregated structures for PHE like in all-atom simulations right?
Any guidance on this query will be hugely helpful.
Thanks in advance
Please Log in or Create an account to join the conversation.
- riccardo
-
- Offline
- Platinum Boarder
3 years 7 months ago #8692
by riccardo
Replied by riccardo on topic Martini studies on aggregation of Amino acids in Lipid bilayer
No Martini studies of simple amino acids aggregating in bilayers comes to my mind, but I would guess there must be something. Translocation PMFs of amino acids (which related to what you are interested in) were computed for example in
doi.org/10.1021/ct300646g
.
I would guess PHE aggregates in bulk water, based on what PHE-PHE does. Indeed, maybe a nice reference to start with just slightly more complex peptides (dipeptides) is this one: doi.org/10.1021/jz2010573 .
I would guess PHE aggregates in bulk water, based on what PHE-PHE does. Indeed, maybe a nice reference to start with just slightly more complex peptides (dipeptides) is this one: doi.org/10.1021/jz2010573 .
Please Log in or Create an account to join the conversation.
- shakira
-
Topic Author
- Offline
- Junior Boarder
Less
More
- Posts: 21
3 years 7 months ago #8695
by shakira
Replied by shakira on topic Martini studies on aggregation of Amino acids in Lipid bilayer
Thanks for the reply. I was trying to reproduce the aggregation of dipeptide (Diphenylalanine) as mentioned in the above mentioned paper, but could not see such aggregation for my MARTINI system.
I am using the martinize.py script to generate the starting structure and topology for diphenylalanine from its all atom pdb file generated from Avogadro. I guess this is the normal procedure to generate the starting files for peptide simulations which I have understood by following the tutorials on peptide in the MARTINI page. Then where am I going wrong?
Can anyone help me with this problem?
Thanks in advance
I am using the martinize.py script to generate the starting structure and topology for diphenylalanine from its all atom pdb file generated from Avogadro. I guess this is the normal procedure to generate the starting files for peptide simulations which I have understood by following the tutorials on peptide in the MARTINI page. Then where am I going wrong?
Can anyone help me with this problem?
Thanks in advance
Please Log in or Create an account to join the conversation.
- siewert
-
- Offline
- Admin
Less
More
- Posts: 61
3 years 7 months ago #8696
by siewert
Replied by siewert on topic Martini studies on aggregation of Amino acids in Lipid bilayer
To understand what is going wrong, we need to know whether you are using just the amino acid side chain (or analogs), or are you including the peptide backbone ? And in case of the latter, do you keep the termini charged or neutral ? And does this correspond to the experiments you want to mimic ?
Please Log in or Create an account to join the conversation.
- shakira
-
Topic Author
- Offline
- Junior Boarder
Less
More
- Posts: 21
3 years 7 months ago #8697
by shakira
Replied by shakira on topic Martini studies on aggregation of Amino acids in Lipid bilayer
We are using the zwitter ionic form of the dipeptide corresponding to the experiment we are trying to mimic. The genearted itp file for dipeptide is pasted below.
; MARTINI (martini22) Coarse Grained topology file for "Protein_A"
; Created by py version 2.6
; Using the following options: -f F-F.pdb -o NEW.top -x NEW-CG.pdb -dssp ./dssp -p backbone -ff martini22
; Sequence:
; FF
; Secondary Structure:
; CC
[ moleculetype ]
; Name Exclusions
Protein_A 1
[ atoms ]
1 Qd 1 PHE BB 1 1.0000 ; C
2 SC5 1 PHE SC1 2 0.0000 ; C
3 SC5 1 PHE SC2 3 0.0000 ; C
4 SC5 1 PHE SC3 4 0.0000 ; C
5 Qa 2 PHE BB 5 -1.0000 ; C
6 SC5 2 PHE SC1 6 0.0000 ; C
7 SC5 2 PHE SC2 7 0.0000 ; C
8 SC5 2 PHE SC3 8 0.0000 ; C
[ bonds ]
; Backbone bonds
1 5 1 0.35000 1250 ; PHE(C)-PHE(C)
; Sidechain bonds
1 2 1 0.31000 7500 ; PHE
5 6 1 0.31000 7500 ; PHE
[ constraints ]
2 3 1 0.27000 ; PHE
2 4 1 0.27000 ; PHE
3 4 1 0.27000 ; PHE
6 7 1 0.27000 ; PHE
6 8 1 0.27000 ; PHE
7 8 1 0.27000 ; PHE
[ angles ]
; Backbone angles
; Backbone-sidechain angles
2 1 5 2 100 25 ; PHE(C)-PHE(C) SBB
1 5 6 2 100 25 ; PHE(C)-PHE(C) SBB
; Sidechain angles
1 2 3 2 150 50 ; PHE
1 2 4 2 150 50 ; PHE
5 6 7 2 150 50 ; PHE
5 6 8 2 150 50 ; PHE
[ dihedrals ]
; Backbone dihedrals
; Sidechain improper dihedrals
1 3 4 2 2 0 50 ; PHE
5 7 8 6 2 0 50 ; PHE
#ifdef POSRES
#ifndef POSRES_FC
#define POSRES_FC 1000.00
#endif
[ position_restraints ]
1 1 POSRES_FC POSRES_FC POSRES_FC
5 1 POSRES_FC POSRES_FC POSRES_FC
#endif
; MARTINI (martini22) Coarse Grained topology file for "Protein_A"
; Created by py version 2.6
; Using the following options: -f F-F.pdb -o NEW.top -x NEW-CG.pdb -dssp ./dssp -p backbone -ff martini22
; Sequence:
; FF
; Secondary Structure:
; CC
[ moleculetype ]
; Name Exclusions
Protein_A 1
[ atoms ]
1 Qd 1 PHE BB 1 1.0000 ; C
2 SC5 1 PHE SC1 2 0.0000 ; C
3 SC5 1 PHE SC2 3 0.0000 ; C
4 SC5 1 PHE SC3 4 0.0000 ; C
5 Qa 2 PHE BB 5 -1.0000 ; C
6 SC5 2 PHE SC1 6 0.0000 ; C
7 SC5 2 PHE SC2 7 0.0000 ; C
8 SC5 2 PHE SC3 8 0.0000 ; C
[ bonds ]
; Backbone bonds
1 5 1 0.35000 1250 ; PHE(C)-PHE(C)
; Sidechain bonds
1 2 1 0.31000 7500 ; PHE
5 6 1 0.31000 7500 ; PHE
[ constraints ]
2 3 1 0.27000 ; PHE
2 4 1 0.27000 ; PHE
3 4 1 0.27000 ; PHE
6 7 1 0.27000 ; PHE
6 8 1 0.27000 ; PHE
7 8 1 0.27000 ; PHE
[ angles ]
; Backbone angles
; Backbone-sidechain angles
2 1 5 2 100 25 ; PHE(C)-PHE(C) SBB
1 5 6 2 100 25 ; PHE(C)-PHE(C) SBB
; Sidechain angles
1 2 3 2 150 50 ; PHE
1 2 4 2 150 50 ; PHE
5 6 7 2 150 50 ; PHE
5 6 8 2 150 50 ; PHE
[ dihedrals ]
; Backbone dihedrals
; Sidechain improper dihedrals
1 3 4 2 2 0 50 ; PHE
5 7 8 6 2 0 50 ; PHE
#ifdef POSRES
#ifndef POSRES_FC
#define POSRES_FC 1000.00
#endif
[ position_restraints ]
1 1 POSRES_FC POSRES_FC POSRES_FC
5 1 POSRES_FC POSRES_FC POSRES_FC
#endif
Please Log in or Create an account to join the conversation.
- shakira
-
Topic Author
- Offline
- Junior Boarder
Less
More
- Posts: 21
3 years 7 months ago #8698
by shakira
Replied by shakira on topic Martini studies on aggregation of Amino acids in Lipid bilayer
Hi all
I have understood the issue with dipeptide system aggregation. I was using the updated force field (version 2.2) for amino acids which does not work!!!! The older version 2.1 shows dipeptide aggregation of diphenylalanine in bulk water. however if you use the updated version you wont be able to see such aggregation as happened in my case.
I have understood the issue with dipeptide system aggregation. I was using the updated force field (version 2.2) for amino acids which does not work!!!! The older version 2.1 shows dipeptide aggregation of diphenylalanine in bulk water. however if you use the updated version you wont be able to see such aggregation as happened in my case.
Please Log in or Create an account to join the conversation.
- riccardo
-
- Offline
- Platinum Boarder
3 years 7 months ago #8703
by riccardo
Replied by riccardo on topic Martini studies on aggregation of Amino acids in Lipid bilayer
Thanks for following up. The only thing that should differ in this case is the SC5 bead of PHE being SC4 in Martini 2.1. Surely SC4 is more hydrophobic than SC5 (which would mean increased aggregation in water) but I would expect PHE-PHE to aggregate also in Martini 2.2. What's the behavior of Martini 2.2? PHE-PHE molecules remain completely soluble or they form smaller aggregates? Did you check/visualize the trajectories?
Please Log in or Create an account to join the conversation.
- shakira
-
Topic Author
- Offline
- Junior Boarder
Less
More
- Posts: 21
3 years 7 months ago #8704
by shakira
Replied by shakira on topic Martini studies on aggregation of Amino acids in Lipid bilayer
Hi
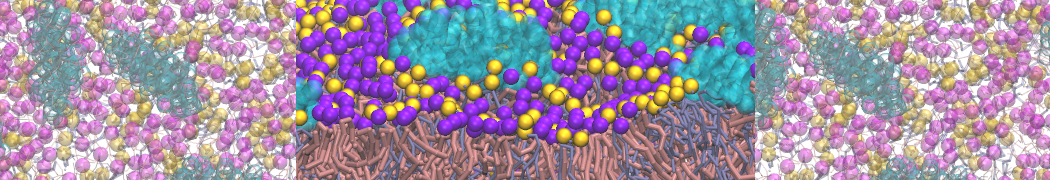
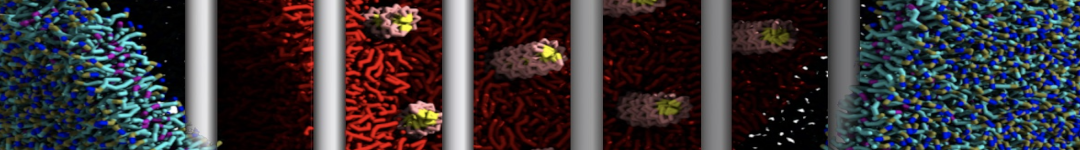
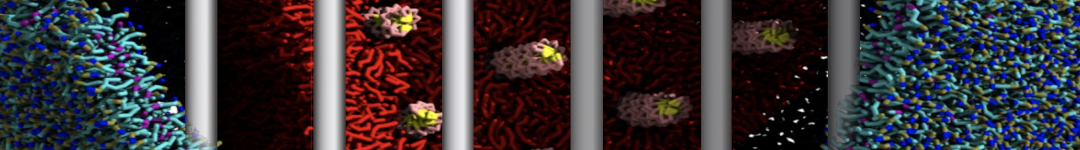
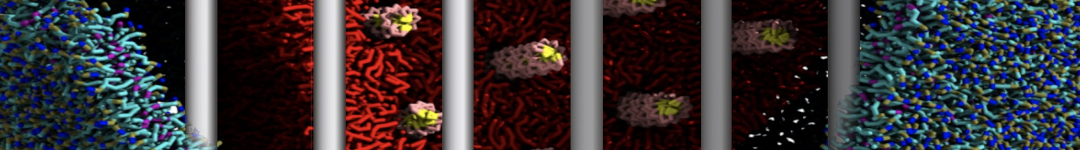
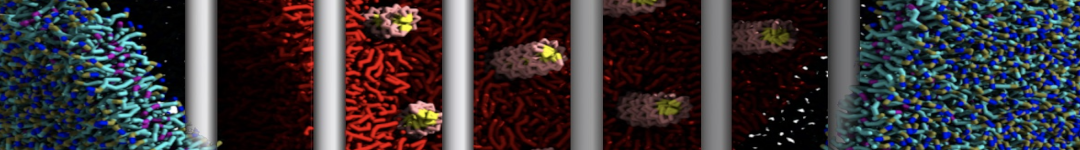
Yes we do checked PHE-PHE aggregation in bulk water with Martini 2.1. It do not show clustering unlike its dipeptide. I have ran the trajectory for 3 microsecond and visualized them in vmd. From the initial random arrangement of PHE molecules some of them grouped together. However it did not show patterns like vesicles, nanotubes or bilayer structures as seen for diphenylalanine. So Martini 2.1 with SC4 particles type also did not improved the results for PHE-PHE aggregation. Probably the limitations of CG water bead drives this.
Yes we do checked PHE-PHE aggregation in bulk water with Martini 2.1. It do not show clustering unlike its dipeptide. I have ran the trajectory for 3 microsecond and visualized them in vmd. From the initial random arrangement of PHE molecules some of them grouped together. However it did not show patterns like vesicles, nanotubes or bilayer structures as seen for diphenylalanine. So Martini 2.1 with SC4 particles type also did not improved the results for PHE-PHE aggregation. Probably the limitations of CG water bead drives this.
Please Log in or Create an account to join the conversation.
- riccardo
-
- Offline
- Platinum Boarder
3 years 7 months ago #8705
by riccardo
Replied by riccardo on topic Martini studies on aggregation of Amino acids in Lipid bilayer
Alright. Sorry, perhaps my post wasn't clear but I meant that I found strange that diphenylalanine doesn't aggregate in Martini 2.2. So my question was: what happens in the trajectories of diphenylalanine in water with Martini 2.2?
For the single phenylalanine, as Siewert asked: are you including the peptide backbone? I can see your diphenylalanine itp but not the phenylalanine one.
For the single phenylalanine, as Siewert asked: are you including the peptide backbone? I can see your diphenylalanine itp but not the phenylalanine one.
Please Log in or Create an account to join the conversation.
- shakira
-
Topic Author
- Offline
- Junior Boarder
Less
More
- Posts: 21
3 years 7 months ago #8706
by shakira
Replied by shakira on topic Martini studies on aggregation of Amino acids in Lipid bilayer
With Martini 2.2 no aggregation was observed neither for phenylalanine nor for diphenylalanine even after 5 microsecond simulation runs. Yes its a bit strange since even with Martini 2.2 the hydrophobicity is well expressed enough for the aggregation.
For single phenylalanine I have used the martinize.py script to generate the CG structure and included the following itp for the aminoacid obtained from Martini downloads for aminoacids (martini_v2.2_aminoacids.itp)
;;; PHENYLALANINE
[ moleculetype ]
; molname nrexcl
PHE 1
[ atoms ]
;id type resnr residu atom cgnr charge
1 P5 1 PHE BAS 1 0
2 SC5 1 PHE SI1 2 0 ; three side chains in triangle
3 SC5 1 PHE SI2 3 0 ; configuration, mimicking
4 SC5 1 PHE SI3 4 0 ; ring structure
[bonds]
; i j funct length force.c.
1 2 1 0.31 7500
[constraints]
; i j funct length
2 3 1 0.27
2 4 1 0.27
3 4 1 0.27
[angles]
; i j k funct angle force.c.
1 2 3 2 150.000 50.0
1 2 4 2 150.000 50.0
[dihedrals]
; i j k l funct angle force.c.
1 3 4 2 2 0.0 50.0 ; to prevent backflipping of ring
For single phenylalanine I have used the martinize.py script to generate the CG structure and included the following itp for the aminoacid obtained from Martini downloads for aminoacids (martini_v2.2_aminoacids.itp)
;;; PHENYLALANINE
[ moleculetype ]
; molname nrexcl
PHE 1
[ atoms ]
;id type resnr residu atom cgnr charge
1 P5 1 PHE BAS 1 0
2 SC5 1 PHE SI1 2 0 ; three side chains in triangle
3 SC5 1 PHE SI2 3 0 ; configuration, mimicking
4 SC5 1 PHE SI3 4 0 ; ring structure
[bonds]
; i j funct length force.c.
1 2 1 0.31 7500
[constraints]
; i j funct length
2 3 1 0.27
2 4 1 0.27
3 4 1 0.27
[angles]
; i j k funct angle force.c.
1 2 3 2 150.000 50.0
1 2 4 2 150.000 50.0
[dihedrals]
; i j k l funct angle force.c.
1 3 4 2 2 0.0 50.0 ; to prevent backflipping of ring
Please Log in or Create an account to join the conversation.
- peterkroon
-
- Offline
- Gold Boarder
Less
More
- Posts: 210
3 years 7 months ago #8707
by peterkroon
Replied by peterkroon on topic Martini studies on aggregation of Amino acids in Lipid bilayer
Hi Shakira,
how do you treat the termini? The parameters in martini_v2.2_aminoacids.itp are intended for non-terminal amino acids.
how do you treat the termini? The parameters in martini_v2.2_aminoacids.itp are intended for non-terminal amino acids.
Please Log in or Create an account to join the conversation.
- riccardo
-
- Offline
- Platinum Boarder
3 years 7 months ago - 3 years 7 months ago #8708
by riccardo
Replied by riccardo on topic Martini studies on aggregation of Amino acids in Lipid bilayer
Alright, thanks! So you have the backbone bead as P5, which I think it's fine. (but Peter knows more!). I have no direct experience so I was a bit curious.
But so you do not observe even small aggregates which perhaps are stable only for 10s-100s of ns for diphenylalanine with Martini 2.2?
But so you do not observe even small aggregates which perhaps are stable only for 10s-100s of ns for diphenylalanine with Martini 2.2?
Last edit: 3 years 7 months ago by riccardo.
Please Log in or Create an account to join the conversation.
- peterkroon
-
- Offline
- Gold Boarder
Less
More
- Posts: 210
3 years 7 months ago #8710
by peterkroon
Replied by peterkroon on topic Martini studies on aggregation of Amino acids in Lipid bilayer
Zwitterionic amino acids are always a bit funny. I'm not 100% sure they've been studied with Martini to begin with.
Just to cover all the bases: There is water in the simulation, right? And could you post your mdp parameters?
Just to cover all the bases: There is water in the simulation, right? And could you post your mdp parameters?
Please Log in or Create an account to join the conversation.
- shakira
-
Topic Author
- Offline
- Junior Boarder
Less
More
- Posts: 21
3 years 7 months ago #8711
by shakira
Replied by shakira on topic Martini studies on aggregation of Amino acids in Lipid bilayer
Hi Peterkroon
I am new to this proteins and peptides section of Martini systems. The PHE aminoacid is represented by 4 beads in Martini right? So I have used it including the martini_v2.2_aminoacids.itp file in the topology. What else should I have done?
For diphenylalanine I used martinize.py script to generate the peptide.itp file as pasted in the previous post.
What was wrong with my approach? Any help will be highly appreciated
I am new to this proteins and peptides section of Martini systems. The PHE aminoacid is represented by 4 beads in Martini right? So I have used it including the martini_v2.2_aminoacids.itp file in the topology. What else should I have done?
For diphenylalanine I used martinize.py script to generate the peptide.itp file as pasted in the previous post.
What was wrong with my approach? Any help will be highly appreciated
Please Log in or Create an account to join the conversation.
- shakira
-
Topic Author
- Offline
- Junior Boarder
Less
More
- Posts: 21
3 years 7 months ago #8712
by shakira
Replied by shakira on topic Martini studies on aggregation of Amino acids in Lipid bilayer
Hi
The mdp parameters I have been using is as follows:
; RUN CONTROL PARAMETERS =
; MARTINI - Most simulations are stable with dt=40 fs,
; some (especially rings) require 20-30 fs.
; The range of time steps used for parametrization
; is 20-40 fs, using smaller time steps is therefore not recommended.
integrator = md
; start time and timestep in ps
tinit = 0.0
dt = 0.025
nsteps = 500000
; number of steps for center of mass motion removal =
nstcomm = 100
comm-grps =
; OUTPUT CONTROL OPTIONS =
; Output frequency for coords (x), velocities (v) and forces (f) =
nstxout = 0
nstvout = 0
nstfout = 0
; Output frequency for energies to log file and energy file =
nstlog = 5000
nstenergy = 5000
; Output frequency and precision for xtc file =
nstxout-compressed = 5000
compressed-x-precision = 100
; This selects the subset of atoms for the xtc file. You can =
; select multiple groups. By default all atoms will be written. =
xtc-grps =
; Selection of energy groups =
energygrps =
; NEIGHBORSEARCHING PARAMETERS =
; MARTINI - no need for more frequent updates
; or larger neighborlist cut-off due
; to the use of shifted potential energy functions.
cutoff-scheme = Verlet
; nblist update frequency =
nstlist = 20
; ns algorithm (simple or grid) =
ns_type = grid
; Periodic boundary conditions: xyz or no =
pbc = xyz
; nblist cut-off =
rlist = 1.1
verlet-buffer-tolerance = 0.05
; OPTIONS FOR ELECTROSTATICS AND VDW =
; MARTINI - vdw and electrostatic interactions are used
; in their shifted forms. Changing to other types of
; electrostatics will affect the general performance of
; the model.
; Method for doing electrostatics =
coulombtype = reaction-field
rcoulomb = 1.1
; Dielectric constant (DC) for cut-off or DC of reaction field =
epsilon_r = 15
epsilon_rf = 0
; Method for doing Van der Waals =
vdw_type = cut-off
; cut-off lengths =
rvdw = 1.1
vdw-modifier = Potential-shift-verlet
; Apply long range dispersion corrections for Energy and Pressure =
DispCorr = No
; OPTIONS FOR WEAK COUPLING ALGORITHMS =
; MARTINI - normal temperature and pressure coupling schemes
; can be used. It is recommended to couple individual groups
; in your system seperately.
; Temperature coupling
tcoupl = Berendsen
tau_t = 0.3 0.3
tc-grps = protein non-protein
ref_t = 303 303
Pcoupl = Berendsen
Pcoupltype = isotropic
tau_p = 3.0
compressibility = 3e-4
ref_p = 1.0
; GENERATE VELOCITIES FOR STARTUP RUN =
gen_vel = yes
gen_temp = 303
gen_seed = 1234
; OPTIONS FOR BONDS =
; MARTINI - for ring systems constraints are defined
; which are best handled using Lincs.
constraints = none
; Type of constraint algorithm
constraint_algorithm = Lincs
; Do not constrain the start configuration =
continuation = no
; Highest order in the expansion of the constraint coupling matrix =
lincs_order = 4
; Lincs will write a warning to the stderr if in one step a bond =
; rotates over more degrees than =
lincs_warnangle = 30
The mdp parameters I have been using is as follows:
; RUN CONTROL PARAMETERS =
; MARTINI - Most simulations are stable with dt=40 fs,
; some (especially rings) require 20-30 fs.
; The range of time steps used for parametrization
; is 20-40 fs, using smaller time steps is therefore not recommended.
integrator = md
; start time and timestep in ps
tinit = 0.0
dt = 0.025
nsteps = 500000
; number of steps for center of mass motion removal =
nstcomm = 100
comm-grps =
; OUTPUT CONTROL OPTIONS =
; Output frequency for coords (x), velocities (v) and forces (f) =
nstxout = 0
nstvout = 0
nstfout = 0
; Output frequency for energies to log file and energy file =
nstlog = 5000
nstenergy = 5000
; Output frequency and precision for xtc file =
nstxout-compressed = 5000
compressed-x-precision = 100
; This selects the subset of atoms for the xtc file. You can =
; select multiple groups. By default all atoms will be written. =
xtc-grps =
; Selection of energy groups =
energygrps =
; NEIGHBORSEARCHING PARAMETERS =
; MARTINI - no need for more frequent updates
; or larger neighborlist cut-off due
; to the use of shifted potential energy functions.
cutoff-scheme = Verlet
; nblist update frequency =
nstlist = 20
; ns algorithm (simple or grid) =
ns_type = grid
; Periodic boundary conditions: xyz or no =
pbc = xyz
; nblist cut-off =
rlist = 1.1
verlet-buffer-tolerance = 0.05
; OPTIONS FOR ELECTROSTATICS AND VDW =
; MARTINI - vdw and electrostatic interactions are used
; in their shifted forms. Changing to other types of
; electrostatics will affect the general performance of
; the model.
; Method for doing electrostatics =
coulombtype = reaction-field
rcoulomb = 1.1
; Dielectric constant (DC) for cut-off or DC of reaction field =
epsilon_r = 15
epsilon_rf = 0
; Method for doing Van der Waals =
vdw_type = cut-off
; cut-off lengths =
rvdw = 1.1
vdw-modifier = Potential-shift-verlet
; Apply long range dispersion corrections for Energy and Pressure =
DispCorr = No
; OPTIONS FOR WEAK COUPLING ALGORITHMS =
; MARTINI - normal temperature and pressure coupling schemes
; can be used. It is recommended to couple individual groups
; in your system seperately.
; Temperature coupling
tcoupl = Berendsen
tau_t = 0.3 0.3
tc-grps = protein non-protein
ref_t = 303 303
Pcoupl = Berendsen
Pcoupltype = isotropic
tau_p = 3.0
compressibility = 3e-4
ref_p = 1.0
; GENERATE VELOCITIES FOR STARTUP RUN =
gen_vel = yes
gen_temp = 303
gen_seed = 1234
; OPTIONS FOR BONDS =
; MARTINI - for ring systems constraints are defined
; which are best handled using Lincs.
constraints = none
; Type of constraint algorithm
constraint_algorithm = Lincs
; Do not constrain the start configuration =
continuation = no
; Highest order in the expansion of the constraint coupling matrix =
lincs_order = 4
; Lincs will write a warning to the stderr if in one step a bond =
; rotates over more degrees than =
lincs_warnangle = 30
Please Log in or Create an account to join the conversation.
- shakira
-
Topic Author
- Offline
- Junior Boarder
Less
More
- Posts: 21
3 years 7 months ago #8713
by shakira
Replied by shakira on topic Martini studies on aggregation of Amino acids in Lipid bilayer
For the zwitter ionic form of the dipeptide the termini is taken ad Qd and Qa particle type in the itp file.
Please Log in or Create an account to join the conversation.
- peterkroon
-
- Offline
- Gold Boarder
Less
More
- Posts: 210
3 years 7 months ago #8714
by peterkroon
Replied by peterkroon on topic Martini studies on aggregation of Amino acids in Lipid bilayer
Your MDP looks mostly sane, don't use two tcoupl groups if there is enough energy exchange between them (as is the case with dissolved (di)peptides).
For the dipeptide I'm pretty sure your itp is correct. I'm not 100% sure on the single peptide, but I don't know what else it should be.
What's the concentration of peptides in your simulation? It could be that at the concentration you use they shouldn't aggregate. Are you sure you don't see short-lived aggregates? Could you post your top file?
If you haven't already, also take a look at [1] where the aggregation of tripeptides is studied in Martini 2.2.
Same for [2], where PHE is reparametrized for Martini 2.2.
If nothing works, consider calculating a dimerization PMF for PHE.
[1] P.W.J.M. Frederix, G.G. Scott, Y.M. Abul-Haija, D. Kalafatovic, C.G. Pappas, N. Javid, et al., Exploring the sequence space for (tri-)peptide self-assembly to design and discover new hydrogels, Nat. Chem. 7 (2015) 30–37. doi:10.1038/nchem.2122.
[2] D.H. de Jong, G. Singh, W.F.D. Bennett, C. Arnarez, T. a Wassenaar, L. V. Schäfer, et al., Improved Parameters for the Martini Coarse-Grained Protein Force Field, J. Chem. Theory Comput. 9 (2013) 687–697. doi:10.1021/ct300646g.
For the dipeptide I'm pretty sure your itp is correct. I'm not 100% sure on the single peptide, but I don't know what else it should be.
What's the concentration of peptides in your simulation? It could be that at the concentration you use they shouldn't aggregate. Are you sure you don't see short-lived aggregates? Could you post your top file?
If you haven't already, also take a look at [1] where the aggregation of tripeptides is studied in Martini 2.2.
Same for [2], where PHE is reparametrized for Martini 2.2.
If nothing works, consider calculating a dimerization PMF for PHE.
[1] P.W.J.M. Frederix, G.G. Scott, Y.M. Abul-Haija, D. Kalafatovic, C.G. Pappas, N. Javid, et al., Exploring the sequence space for (tri-)peptide self-assembly to design and discover new hydrogels, Nat. Chem. 7 (2015) 30–37. doi:10.1038/nchem.2122.
[2] D.H. de Jong, G. Singh, W.F.D. Bennett, C. Arnarez, T. a Wassenaar, L. V. Schäfer, et al., Improved Parameters for the Martini Coarse-Grained Protein Force Field, J. Chem. Theory Comput. 9 (2013) 687–697. doi:10.1021/ct300646g.
Please Log in or Create an account to join the conversation.
Time to create page: 0.122 seconds